
By Vince Giuliano and James P Watson.
Trehalose is a natural disaccharide sugar contained in mushrooms and many other organisms. It has remarkable health-producing and life-extending properties that work in mice and lower organisms. That is why we choose to focus on it here. This blog entry details animal model research which suggests significant health and longevity advantages of trehalose consumption. We describe issues related to its bioavailability and safety in humans and wonder why it is not in clinical trials for neurodegenerative diseases.
Basics About trehalose:
“In nature, Trehaose can be found in animals, plants, and microorganisms. In animals, trehalose is prevalent in shrimp, and also in insects, including grasshoppers, locusts, butterflies, and bees, in which blood-sugar is trehalose. The trehalose is then broken down into glucose by the catabolic enzyme trehalase for use. Trehalose is also present in the nutrition exchange liquid of hornets and their larvae(ref).” This is unlike the situation in us humans where our primary blood sugar is glucose. When we eat trehalose, some or much of it is broken down into glucose by the trehalase enzyme in the gut before it enters our bloodstreams.
The enzyme trehalase converts trehalose to glucose in humans and most animals
“In general, the fate of ingested or parenterally administered trehalose corresponds to that of glucose since trehalose is rapidly hydrolysed to glucose by the enzyme trehalase. Trehalase is found in humans and most animals at the brush border of the intestinal mucosa, as well as in the kidney, liver, and blood plasma (Hore & Messer, 1968; van Handel, 1970; Demelier et al., 1975; Labat-Robert, 1982; Niederst & Dauça, 1985; Eze, 1989; Riby et al., 1990; Yoshida, 1993). Trehalase activity has been found in the small intestine of humans, mice, rats, guinea-pigs, rabbits, pigs, and baboons (Cerda et al., 1972; Hietanen, 1973; Ruppin et al., 1974; Maestracci, 1976; Garland, 1989)(from WHO FOOD ADDITIVES SERIES 46:TREHALOSE). The implication is that conversion of trehalose to glucose starts in the gut and normally is continued and finished elsewhere. I am not clear how much of trehalose is converted where under what circumstances. In animal experiments to be described, ingested trehalase has impacts on cells quite opposite of those of glucose impacts.
Trehalose is safe and non-toxic for humans – when eaten
It has been used as a food sweetener in Japan for many years. A January article in Food Chemistry showed that in mice, doses of 1.25gms/kg, 2.5gms/kg, and 5gms/kg were safe Multiple toxicity studies of trehalose in mice by intragastric administration: “The results obtained in the present study could support the conclusion that consumption of trehalose has no adverse effects for humans.” This article suggests that as much as 5gms per day is safe in humans: Trehalose: a review of properties, history of use and human tolerance, and results of multiple safety studies reports: “No consistent treatment-related, dose-dependent adverse effects were observed in any of the eight safety studies performed at doses up to 10% of the diets. On the basis of these toxicity studies, human studies in which doses of trehalose were administered to various populations, and consumption of trehalose in commercial products in Japan, it is concluded that trehalose is safe for use as an ingredient in consumer products when used in accordance with current Good Manufacturing Practices.”
We note that non-toxicity is probably a corollary of well-functioning of the trehalse enzyme. Evolution invented trehalase in the gut to convert trehalose into glucose before it enters the blood stream. So, administering trehalose intravenously or via liposomes might or might not be so safe. We discuss this issue further below.
Mushrooms are a great source of Trehalose
They sometimes call trehalose “mushroom sugar.” Here are the main edible mushrooms that have trehalose
- shiitake (Lentinula edodes)
- maitake (Grifola fondosa),
- nameko (Pholiota nameko)
- Judas’s ear (Auricularia auricula-judae)
They can contain 1% to 17% percent of trehalose in dry weight form.
Trehalose stabilizes protein structures
For a long time, trehalose was seen mainly as yet-another sugar occurring in nature. Now. it is known to play very important roles in stabilizing protein structures, the root reason why it is of interest for health and longevity.
The 2009 publication Effect of trehalose on protein structure tells this story: “Trehalose is a ubiquitous molecule that occurs in lower and higher life forms but not in mammals. Till about 40 years ago, trehalose was visualized as a storage molecule, aiding the release of glucose for carrying out cellular functions. This perception has now changed dramatically. The role of trehalose has expanded, and this molecule has now been implicated in a variety of situations. Trehalose is synthesized as a stress-responsive factor when cells are exposed to environmental stresses like heat, cold, oxidation, desiccation, and so forth. When unicellular organisms are exposed to stress, they adapt by synthesizing huge amounts of trehalose, which helps them in retaining cellular integrity. This is thought to occur by prevention of denaturation of proteins by trehalose, which would otherwise degrade under stress. This explanation may be rational, since recently, trehalose has been shown to slow down the rate of polyglutamine-mediated protein aggregation and the resultant pathogenesis by stabilizing an aggregation-prone model protein. In recent years, trehalose has also proved useful in the cryopreservation of sperm and stem cells and in the development of a highly reliable organ preservation solution. This review aims to highlight the changing perception of the role of trehalose over the last 10 years and to propose common mechanisms that may be involved in all the myriad ways in which trehalose stabilizes protein structures. These will take into account the structure of trehalose molecule and its interactions with its environment, and the explanations will focus on the role of trehalose in preventing protein denaturation.” There is much continuing research on trehalose since 2009. Pubmed.org lists 10,428 research publication citations for trehalose
 Hayashibara and Cargill have made a food sweetener product out of Trehalose called TREHA, and they received approval last year for it to be marketed in Canada. It has a relative sweetness of 40-45%, compared to sucrose. Hayashibara makes this in vats from food-grade starch. A one-pound jar of “pure trehalose” can be purchased online for $9.99(ref).
Hayashibara and Cargill have made a food sweetener product out of Trehalose called TREHA, and they received approval last year for it to be marketed in Canada. It has a relative sweetness of 40-45%, compared to sucrose. Hayashibara makes this in vats from food-grade starch. A one-pound jar of “pure trehalose” can be purchased online for $9.99(ref).
Research findings make trehalose very interesting
A substantial portion of the research literature on trehalose relates primarily to plants, chemical relationships, food preservation and industrial processes. We focus here on items related to health.
1. Trehalose enhances longevity by inhibiting the Insulin/IGF axis in nematodes.
The IGF1 pathway activated by insulin is know to be associated with accelerated aging and age-related degenerative diseases, and is frequently activated in older people, though it plays important roles in all stages of human development. In fact we believe that activation of this pathway is a root cause of aging. The 2010 publication Trehalose extends longevity in the nematode Caenorhabditis elegans reports: “Trehalose is a disaccharide of glucose found in diverse organisms and is suggested to act as a stress protectant against heat, cold, desiccation, anoxia, and oxidation. Here, we demonstrate that treatment of Caenorhabditis elegans with trehalose starting from the young-adult stage extended the mean life span by over 30% without any side effects. Surprisingly, trehalose treatment starting even from the old-adult stage shortly thereafter retarded the age-associated decline in survivorship and extended the remaining life span by 60%. Demographic analyses of age-specific mortality rates revealed that trehalose extended the life span by lowering age-independent vulnerability. Moreover, trehalose increased the reproductive span and retarded the age-associated decrease in pharyngeal-pumping rate and the accumulation of lipofuscin autofluorescence. Trehalose also enhanced thermotolerance and reduced polyglutamine aggregation. These results suggest that trehalose suppressed aging by counteracting internal or external stresses that disrupt protein homeostasis. On the other hand, the life span-extending effect of trehalose was abolished in long-lived insulin/IGF-1-like receptor (daf-2) mutants. RNA interference-mediated inactivation of the trehalose-biosynthesis genes trehalose-6-phosphate synthase-1 (tps-1) and tps-2, which are known to be up-regulated in daf-2 mutants, decreased the daf-2 life span. These findings indicate that a reduction in insulin/IGF-1-like signaling extends life span, at least in part, through the aging-suppressor function of trehalose. Trehalose may be a lead compound for potential nutraceutical intervention of the aging process.”
Although other interventions in nematodes can do more, 60% life extension in older animals is not too shabby and led us to wonder further about exactly how trehalose works.
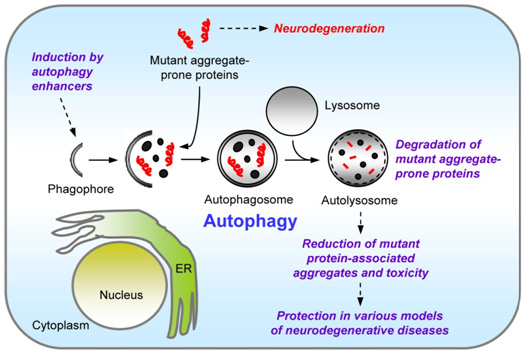
2. Trehalose induces autophagy independently of mTOR pathway inhibition and has been shown in experimental models to help in neurodegenerative diseases. Trehalose can clear alpha-synuclein, polyglutamate, amyloid beta 40, amyloid beta 42, and tau proteins such as this seen in Huntington’s disease, Parkinson’s disease, and Alzheimer’s disease.
A series of publications going back to 2004 establish these important points.
The 2004 publication Trehalose alleviates polyglutamine-mediated pathology in a mouse model of Huntington disease suggested that the beneficial actions of trehalose in neurodegenerative diseases could be as a chaperone molecule affecting protein folding. It reported: “Inhibition of polyglutamine-induced protein aggregation could provide treatment options for polyglutamine diseases such as Huntington disease. Here we showed through in vitro screening studies that various disaccharides can inhibit polyglutamine-mediated protein aggregation. We also found that various disaccharides reduced polyglutamine aggregates and increased survival in a cellular model of Huntington disease. Oral administration of trehalose, the most effective of these disaccharides, decreased polyglutamine aggregates in cerebrum and liver, improved motor dysfunction and extended lifespan in a transgenic mouse model of Huntington disease. We suggest that these beneficial effects are the result of trehalose binding to expanded polyglutamines and stabilizing the partially unfolded polyglutamine-containing protein. Lack of toxicity and high solubility, coupled with efficacy upon oral administration, make trehalose promising as a therapeutic drug or lead compound for the treatment of polyglutamine diseases. The saccharide-polyglutamine interaction identified here thus provides a new therapeutic strategy for polyglutamine diseases.”
In the pancreas, insulin forms a type of amyloid called “amylin.” It is essentially misfolded insulin molecule. Trehalose prevents this from occurring. In this case it may be functioning as a “chemical chaperone,” The 2004 publication Inhibition of insulin amyloid formation by small stress moleculesreported: “Amyloidogenic proteins undergo an alternative folding pathway under stressful conditions leading to formation of fibrils having cross beta-sheet structure, which is the hallmark of many neurodegenerative diseases. As a means of surviving against external stress, on the other hand, many microorganisms accumulate small stress molecules to prevent abnormal protein folding and to contribute to protein stability, which hints at the efficacy of the solutes against amyloid formation. The current work demonstrates the effectiveness of small stress molecules such as ectoine, betaine, trehalose, and citrulline on inhibition of insulin amyloid formation in vitro. The inhibitory effects were analyzed by thioflavin T-induced fluorescence, circular dichroism, and atomic force microscopy. This report suggests that naturally occurring small molecules may serve a function that is typically fulfilled by protein chaperones, and it provides a hint for designing inhibitors against amyloid formation associated with neurodegenerative disorders.
A 2005 publication Trehalose differentially inhibits aggregation and neurotoxicity of beta-amyloid 40 and 42 relates to a possible role for trehalose in preventing or treating Alzheimer’s disease. “These results also suggest that the use of trehalose, a highly soluble, low-priced sugar, as part of a potential therapeutic cocktail to control Abeta peptide aggregation and toxicity warrants further investigation.”
The 2006 publication Trehalose, a Novel mTOR-independent Autophagy Enhancer, Accelerates the Clearance of Mutant Huntingtin and α-Synuclein points out how its autophagic effects are mTOR-independent. “Trehalose, a disaccharide present in many non-mammalian species, protects cells against various environmental stresses. Whereas some of the protective effects may be explained by its chemical chaperone properties, its actions are largely unknown. Here we report a novel function of trehalose as an mTOR-independent autophagy activator. Trehalose-induced autophagy enhanced the clearance of autophagy substrates like mutant huntingtin and the A30P and A53T mutants of α-synuclein, associated with Huntington disease (HD) and Parkinson disease (PD), respectively. Furthermore, trehalose and mTOR inhibition by rapamycin together exerted an additive effect on the clearance of these aggregate-prone proteins because of increased autophagic activity. By inducing autophagy, we showed that trehalose also protects cells against subsequent pro-apoptotic insults via the mitochondrial pathway. The dual protective properties of trehalose (as an inducer of autophagy and chemical chaperone) and the combinatorial strategy with rapamycin may be relevant to the treatment of HD and related diseases, where the mutant proteins are autophagy substrates.”
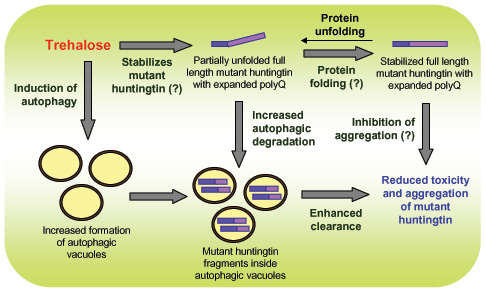
The 2010 publication Trehalose ameliorates dopaminergic and tau pathology in parkin deleted/tau overexpressing mice through autophagy activation reports: “Tauopathies are neurodegenerative diseases, sporadic or familial, mainly characterized by dementia and parkinsonism associated to atrophy of the frontotemporal cortex and the basal ganglia, with deposition of abnormal tau in brain. Hereditary tauopathies are related with mutations of the tau gene. Up to the present, these diseases have not been helped by any disease-modifying treatment, and patients die a few years after the onset of symptoms. — We have developed and characterized a mouse model of tauopathy with parkinsonism, overexpressing human mutated tau protein with deletion of parkin (PK−/−/TauVLW). At 3 months of age, these mice present abnormal dopamine-related behavior, severe dropout of dopamine neurons in the ventral midbrain, reduced dopamine levels in the striatum and abundant phosphorylated tau-positive neuritic plaques, neurofibrillary tangles, astrogliosis, and, at 12 months old, plaques of murine β-amyloid in the hippocampus. — Trehalose is a natural disaccharide that increases the removal of abnormal proteins through enhancement of autophagy. In this work, we tested if 1% trehalose in the drinking water reverts the PK−/−/TauVLWphenotype. The treatment with trehalose of 3-month-old PK−/−/TauVLW mice for 2.5 months reverted the dropout of dopamine neurons, which takes place in the ventral midbrain of vehicle treated PK−/−/TauVLWand the reduced dopamine-related proteins levels in the midbrain and striatum. The number of phosphorylated tau-positive neuritic plaques and the levels of phosphorylated tau decreased, as well as astrogliosis in brain regions. The autophagy markers in the brain, the autophagic vacuoles isolated from the liver, and the electron microscopy data indicate that these effects of trehalose are mediated by autophagy. The treatment with trehalose for 4 months of 3-month-old PK−/−/TauVLW mice maintained the amelioration of the tau pathology and astrogliosis but failed to revert DA-related pathology in the striatum. Furthermore, the 3-week treatment with trehalose of 14-month-old PK−/−/TauVLW mice, at the limit of their life expectancy, improved the motor behavior and anxiety of these animals, and reduced their levels of phosphorylated tau and the number of murine β-amyloid plaques. — Trehalose is neuroprotective in this model of tauopathy. Since trehalose is free of toxic effects at high concentrations, this study opens the way for clinical studies of the effects of trehalose in human tauopathies.”
The 2010 publication Chemical Inducers of Autophagy That Enhance the Clearance of Mutant Proteins in Neurodegenerative Diseases instead suggests that the action of trehalose is via autophagy. It has to say: “Another chemical screen to identify inhibitors of polyQ-mediated protein aggregation in vitro identified t.rehalose (a disaccharide) as an inhibitor of mutant huntingtin aggregation, which reduced toxicity in HD cell models and attenuated disease pathology in a mouse model of HD (59). This protective effect of trehalose was suggested to be mediated by its ability to act as a chemical chaperone through its binding to the polyQ-expanded mutant huntingtin and influencing its protein folding and aggregation. However, we have shown that trehalose enhances the autophagy pathway independently of mTOR, thereby increasing the clearance of mutant aggregate-prone proteins (60). Additionally, trehalose protected against pro-apoptotic insults via autophagy (60). The myriad of protective properties of trehalose acting as an autophagy inducer and chemical chaperone, coupled with its lack of toxicity, may be of benefit in the treatment of neurodegenerative disorders.” The critical importance of autophagy is detailed in our 2013 blog entry Autophagy – the housekeeper in every cell that fights aging.
The 2011 article The accumulation of neurotoxic proteins, induced by proteasome inhibition, is reverted by trehalose, an enhancer of autophagy, in human neuroblastoma cells reports: “Neurodegenerative diseases like Parkinson’s disease, Alzheimer’s disease, Huntington’s disease and others are due to accumulation of abnormal proteins which fold improperly and impair neuronal function. Accumulation of these proteins could be achieved by several mechanisms including mutation, overproduction or impairment of its degradation. Inhibition of the normal protein degradation is produced by blockade of the ubiquitin proteasome system. — We have shown that epoxomicin, a proteasome inhibitor, increases the levels of proteins involved in neurodegenerative disorders such as α-synuclein and hyper phosphorylated tau in NB69 human neuroblastoma cells and that such increase correlates with an enhanced rate of cell death. We then investigated whether the stimulation of autophagy, an alternative mechanism for elimination of abnormal proteins, by treatment with trehalose, counteracts the effects of proteasomal blockade. Trehalose, a disaccharide present in many non-mammalian species, known to enhance autophagy, protects cells against various environmental stresses. Treatment with trehalose produced a dose and time-dependent increase in the number of autophagosomes and markers of autophagy in NB69 cells. Trehalose did not change the number of total neither the number of dividing cells in the culture but it completely prevented the necrosis of NB69 induced by epoxomicin. In addition, the treatment with trehalose reverted the accumulation, induced by epoxomicin, of polyubiquitinated proteins, total and phosphorylated tau, p-GSK-3, and α-synuclein, as well as the α-synuclein intracellular aggregates. The effects of trehalose were not mediated through activation of free radical scavenging compounds, like GSH, or mitochondrial proteins, like DJ1, but trehalose reduced the activation of ERK and chaperone HSP-70 induced by epoxomicin. Inhibition of ERK phosphorylation prevented the epoxomicin-induced cell death. Inhibition of autophagy reverted the neuroprotective effects of trehalose in epoxomicin-induced cell death. These results suggest that trehalose is a powerful modifier of abnormal protein accumulation in neurodegenerative diseases.” Because this experiment was with human cells, the suggestion is that the results observed in mice might well carry over to humans.
The 2012 publication Autophagic degradation of tau in primary neurons and its enhancement by trehalose relates: “Modulating the tau level may represent a therapeutic target for Alzheimer’s disease (AD), as accumulating evidence shows that Abeta-induced neurodegeneration is mediated by tau. It is therefore important to understand the expression and degradation of tau in neurons. Recently we showed that overexpressed mutant tau and tau aggregates are degraded via the autophagic pathway in an N2a cell model. Here we investigated whether autophagy is involved in the degradation of endogenous tau in cultured primary neurons. We activated this pathway in primary neurons with trehalose, an enhancer of autophagy. This resulted in the reduction of endogenous tau protein. Tau phosphorylation at several sites elevated in AD pathology had little influence on its degradation by autophagy. Furthermore, by using a neuronal cell model of tauopathy, we showed that activation of autophagy suppresses tau aggregation and eliminates cytotoxicity. Notably, apart from activating autophagy, trehalose also inhibits tau aggregation directly. Thus, trehalose may be a good candidate for developing therapeutic strategies for AD and other tauopathies.”
Finally, the 2012 publication Stimulation of autophagy reduces neurodegeneration in a mouse model of human tauopathy reports: “The accumulation of insoluble proteins is a pathological hallmark of several neurodegenerative disorders. Tauopathies are caused by the dysfunction and aggregation of tau protein and an impairment of cellular protein degradation pathways may contribute to their pathogenesis. Thus, a deficiency in autophagy can cause neurodegeneration, while activation of autophagy is protective against some proteinopathies. Little is known about the role of autophagy in animal models of human tauopathy. In the present report, we assessed the effects of autophagy stimulation by trehalose in a transgenic mouse model of tauopathy, the human mutant P301S tau mouse, using biochemical and immunohistochemical analyses. Neuronal survival was evaluated by stereology. Autophagy was activated in the brain, where the number of neurons containing tau inclusions was significantly reduced, as was the amount of insoluble tau protein. This reduction in tau aggregates was associated with improved neuronal survival in the cerebral cortex and the brainstem. We also observed a decrease of p62 protein, suggesting that it may contribute to the removal of tau inclusions. Trehalose failed to activate autophagy in the spinal cord, where it had no impact on the level of sarkosyl-insoluble tau. Accordingly, trehalose had no effect on the motor impairment of human mutant P301S tau transgenic mice. Our findings provide direct evidence in favour of the degradation of tau aggregates by autophagy. Activation of autophagy may be worth investigating in the context of therapies for human tauopathies.”
3. Trehalose has also been shown to be effective for treating prion disease.
When prion infected organisms are given trehalose, they start degrading the prions. The response occurred at concentrations of 100 mM – that is not that high!. The 2009 publication Autophagy induction by trehalose counteracts cellular prion infection reports: “Prion diseases are fatal neurodegenerative and infectious disorders for which no therapeutic or prophylactic regimens exist. In search of cellular mechanisms that play a role in prion diseases and have the potential to interfere with accumulation of intracellular pathological prion protein (PrP(Sc)), we investigated the autophagic pathway and one of its recently published inducers, trehalose. Trehalose, an alpha-linked disaccharide, has been shown to accelerate clearance of mutant huntingtin and alpha-synuclein by activating autophagy, mainly in an mTOR-independent manner. Here, we demonstrate that trehalose can significantly reduce PrP(Sc) in a dose- and time-dependent manner while at the same time it induces autophagy in persistently prion-infected neuronal cells. Inhibition of autophagy, either pharmacologically by known autophagy inhibitors like 3-methyladenine, or genetically by siRNA targeting Atg5, counteracted the anti-prion effect of trehalose. Hence, we provide direct experimental evidence that induction of autophagy mediates enhanced cellular degradation of prions. Similar results were obtained with rapamycin, a known inducer of autophagy, and imatinib, which has been shown to activate autophagosome formation. While induction of autophagy resulted in reduction of PrP(Sc), inhibition of autophagy increased the amounts of cellular PrP(Sc), suggesting that autophagy is involved in the physiological degradation process of cellular PrP(Sc). Preliminary in vivo studies with trehalose in intraperitoneally prion-infected mice did not result in prolongation of incubation times, but demonstrated delayed
4. Trehalose can regulate lipid metabolism (Macrolipophagy) via autophagy
The 2010 publication Autophagy regulates lipid metabolism reports “The intracellular storage and utilization of lipids are critical to maintain cellular energy homeostasis. During nutrient deprivation, cellular lipids stored as triglycerides in lipid droplets are hydrolysed into fatty acids for energy. A second cellular response to starvation is the induction of autophagy, which delivers intracellular proteins and organelles sequestered in double-membrane vesicles (autophagosomes) to lysosomes for degradation and use as an energy source. Lipolysis and autophagy share similarities in regulation and function but are not known to be interrelated. Here we show a previously unknown function for autophagy in regulating intracellular lipid stores (macrolipophagy). Lipid droplets and autophagic components associated during nutrient deprivation, and inhibition of autophagy in cultured hepatocytes and mouse liver increased triglyceride storage in lipid droplets. This study identifies a critical function for autophagy in lipid metabolism that could have important implications for human diseases with lipid over-accumulation such as those that comprise the metabolic syndrome.”
We did not previously know about this relationship between autophagy and liquid metabolism. Insulin turns off lipid metabolism and turns on lipid storage. This is why people get fat when their insulin levels are high. Secondly, the cited article points out that when autophagy is inhibited, lipids (triglycerides) built up in the liver. Thus “fatty liver” is due to high insulin levels and autophagy inhibition, NOT dietary fat intake. Then they went on to do an experiment directly linking autophagy to lipid metabolism They called this “macrolipophagy.” I (Jim) think this is why the Atkins diet (or ketogenic diet) works!
5. Illustrative of the versatility and multiple mechanisms of operation of trehalose is its operation in cases of vasospasm.
The 2012 publication Trehalose treatment suppresses inflammation, oxidative stress, and vasospasm induced by experimental subarachnoid hemorrhage reports: “Background: Subarachnoid hemorrhage (SAH) frequently results in several complications, including cerebral vasospasm, associated with high mortality. Although cerebral vasospasm is a major cause of brain damages after SAH, other factors such as inflammatory responses and oxidative stress also contribute to high mortality after SAH. Trehalose is a non-reducing disaccharide in which two glucose units are linked by α,α-1,1-glycosidic bond, and has been shown to induce tolerance to a variety of stressors in numerous organisms. In the present study, we investigated the effect of trehalose on cerebral vasospasm, inflammatory responses, and oxidative stress induced by blood in vitro and in vivo. Methods: Enzyme immunoassay for eicosanoids, pro-inflammatory cytokines, and endothelin-1, and western blotting analysis for cyclooxygenase-2, inducible nitric oxide synthase, and inhibitor of NF-κB were examined in macrophage-like cells treated with hemolysate. After treatment with hemolysate and hydrogen peroxide, the levels of lipid peroxide and amounts of arachidonic acid release were also analyzed. Three hours after the onset of experimental SAH, 18 Japanese White rabbits received an injection of saline, trehalose, or maltose into the cisterna magna. Angiographic and histological analyses of the basilar arteries were performed. In a separate study, the femoral arteries from 60 rats were exposed to fresh autologous blood. At 1, 3, 5, 7, 10, and 20days after treatment, cryosections prepared from the femoral arteries were histologically analyzed. Results: When cells were treated with hemolysate, trehalose inhibited the production of several inflammatory mediators and degradation of the inhibitor of NF-κB and also suppressed the lipid peroxidation, the reactive oxygen species-induced arachidonic acid release in vitro. In the rabbit model, trehalose produced an inhibitory effect on vasospasm after the onset of experimental SAH, while maltose had only a moderate effect. When the rat femoral arteries exposed to blood were investigated for 20days, histological analysis revealed that trehalose suppressed vasospasm, inflammatory response, and lipid peroxidation. Conclusions: These data suggest that trehalose has suppressive effects on several pathological events after SAH, including vasospasm, inflammatory responses, and lipid peroxidation. Trehalose may be a new therapeutic approach for treatment of complications after SAH.”
6. Trehalose penetrates the blood-brain barrier quite well.
Since it has been shown to stimulate autophagy and clear protein aggregates in the brain with neurodegenerative diseases, for trehalose to be effective it would be very important for it to cross the blood brain barrier. It does. This was shown in an Alzheimer’s conference a few years ago. It is an effective treatment for animal models of Alzhiemer’s disease even after the onset of the disease. It is also effective to prevent Alzheimer’s disease from occurring. See the 2010 publication A diverse portfolio of novel drug discovery eff orts for Alzheimer’s disease: Meeting report from the 11th International Conference on Alzheimer’s Drug Discovery, 27–28 September 2010, Jersey City, NJ, USA“Trehalose, which has good blood–brain barrier penetration, was effective as both a therapeutic agent after disease onset as well as a long-term preventative treatment that improved motor function in the JNPL3 mouse model.”
This is more good news.
7. While trehalose is a sugar, it does does not form Advanced Glycation Endproducts (AGEs)
This again was news for us, “Trehalose is a nonreducing sugar formed from two glucose units joined by a 1-1 alpha bond, giving it the name of α-D-glucopyranosyl-(1→1)-α-D-glucopyranoside. The bonding makes trehalose very resistant to acid hydrolysis, and therefore is stable in solution at high temperatures, even under acidic conditions. The bonding also keeps nonreducing sugars in closed-ring form, such that the aldehyde or ketone end groups do not bind to the lysine or arginine residues of proteins (a process called glycation)(ref).” Tissue glycation has been seen as a major problem of aging for a very long time as pointed out in my treatise in 2007(ref). This is yet more good news.
8. Trehalose may prevent the progression of insulin resistance
The 2010 publication Trehalose prevents adipocyte hypertrophy and mitigates insulin resistance reports: “Trehalose has been shown to evoke lower insulin secretion than glucose in oral saccharide tolerance tests in humans. Given this hypoinsulinemic effect of trehalose, we hypothesized that trehalose suppresses adipocyte hypertrophy by reducing storage of triglyceride and mitigates insulin resistance in mice fed a high-fat diet (HFD). Mice were fed an HFD and given drinking water containing 2.5% saccharide (glucose [Glc], trehalose [Tre], maltose [Mal], high-fructose corn syrup, or fructose [Fru]) ad libitum. After 7 weeks of HFD and saccharide intake, fasting serum insulin levels in the Tre/HFD group were significantly lower than in the Mal/HFD and Glc/HFD groups (P < .05). Furthermore, the Tre/HFD group showed a significantly suppressed elevation of homeostasis model assessment-insulin resistance compared with the Mal/HFD group (P < .05) and showed a trend toward lower homeostasis model assessment-insulin resistance than the Glc/HFD group. After 8 weeks of feeding, mesenteric adipocyte size in the Tre/HFD group showed significantly less hypertrophy than the Glc/HFD, Mal/HFD, high-fructose corn syrup/HFD, or Fru/HFD group. Analysis of gene expression in mesenteric adipocytes showed that no statistically significant difference in the expression of monocyte chemoattractant protein-1 (MCP-1) messenger RNA (mRNA) was observed between the Tre/HFD group and the distilled water/standard diet group, whereas a significant increase in the MCP-1 mRNA expression was observed in the Glc/HFD, Mal/HFD, Fru/HFD, and distilled water/HFD groups. Thus, our data indicate that trehalose prevents adipocyte hypertrophy and mitigates insulin resistance in HFD-fed mice by reducing insulin secretion and down-regulating mRNA expression of MCP-1. These findings further suggest that trehalose is a functional saccharide that mitigates insulin resistance.”
The suggestion is that if we could substitute trehalose for conventional sugar, that might go a long way towards solving the diabetes epidemic.
0. Trehalose treatment reverses autophagy impairment and prevents neural tube defects in diabetic pregnancies – in mice at least
The 2013 publication Trehalose prevents neural tube defects by correcting maternal diabetes-suppressed autophagy and neurogenesis reports: “Preexisting maternal diabetes increases the risk of neural tube defects (NTDs). The mechanism underlying maternal diabetes-induced NTDs is not totally defined, and its prevention remains a challenge. Autophagy, an intracellular process to degrade dysfunction protein and damaged cellular organelles, regulates cell proliferation, differentiation, and apoptosis. Because autophagy impairment causes NTDs reminiscent of those observed in diabetic pregnancies, we hypothesize that maternal diabetes-induced autophagy impairment causes NTD formation by disrupting cellular homeostasis, leading to endoplasmic reticulum (ER) stress and apoptosis, and that restoration of autophagy by trehalose, a natural disaccharide, prevents diabetes-induced NTDs. Embryos from nondiabetic and type 1 diabetic mice fed with or without 2 or 5% trehalose water were used to assess markers of autophagy, ER stress, and neurogenesis, numbers of autophagosomes, gene expression that regulates autophagy, NTD rates, indices of mitochondrial dysfunction, and neuroepithelial cell apoptosis. Maternal diabetes suppressed autophagy by significantly reducing LC3-II expression, autophagosome numbers, and GFP-LC3 punctate foci in neuroepithelial cells and by altering autophagy-related gene expression. Maternal diabetes delayed neurogenesis by blocking Sox1 neural progenitor differentiation. Trehalose treatment reversed autophagy impairment and prevented NTDs in diabetic pregnancies. Trehalose resolved homeostatic imbalance by correcting mitochondrial defects, dysfunctional proteins, ER stress, apoptosis, and delayed neurogenesis in the neural tubes exposed to hyperglycemia. Our study demonstrates for the first time that maternal diabetes suppresses autophagy in neuroepithelial cells of the developing neural tube, leading to NTD formation, and provides evidence for the potential efficacy of trehalose as an intervention against hyperglycemia-induced NTDs.”
10. Trehalose exercises an antidepressant affect, at least on mice
The 2013 publication Trehalose induced antidepressant-like effects and autophagy enhancement in mice reports: “RATIONALE: The disaccharide trehalose protects cells from hypoxic and anoxic injury and suppresses protein aggregation. In vivo studies with trehalose show cellular and behavioral beneficial effects in animal models of neurodegenerative diseases. Moreover, trehalose was shown to enhance autophagy, a process that had been recently suggested to be involved in the therapeutic action of antidepressant and mood-stabilizing drugs. OBJECTIVE: The present study was therefore designed to explore antidepressant and mood-stabilizing activity of trehalose in animal models for depression and mania. METHODS: Trehalose 1 or 2% was administered for 3 weeks as a drinking solution to Black Swiss mice (a model of manic-like behaviors) or 2% to ICR mice and their behavior evaluated in a number of tests related to depression or mania. The effects of trehalose were compared with similar chronic administration of the disaccharide maltose as well as with a vehicle (water) control. RESULTS: Chronic administration of trehalose resulted in a reduction of frontal cortex p62/beclin-1 ratio suggesting enhancement of autophagy. Trehalose had no mood-stabilizing effects on manic-like behavior in Black Swiss mice but instead augmented amphetamine-induced hyperactivity, an effect similar to antidepressant drugs. In ICR mice, trehalose did not alter spontaneous activity or amphetamine-induced hyperactivity but in two separate experiments had a significant effect to reduce immobility in the forced swim test, a standard screening test for antidepressant-like effects. CONCLUSIONS: The results suggest that trehalose may have antidepressant-like properties. It is hypothesized that these behavioral changes could be related to trehalose effects to enhance autophagy.”
11. There is an issue of how much trehalose ingested by humans makes it into the bloodstream where it can induce the beneficial effects outlined above.
We mentioned that the small intestine has an enzyme called trehalase. This enzyme is also found in the kidney. The trehalase enzyme breaks the trehalose into two separate glucopyranoside sugar molecules that are then absorbed. One gram of trehalose produces 4 kcals/gm of sugar. From Health Canada archived content: “This disaccharide is enzymatically hydrolysed by the enzyme trehalase in the small intestine into two glucose subunits which are subsequently absorbed and metabolized in a manner similar to maltose. Common disaccharides such as sucrose and lactose are digested through the same physiological process. The sugar provides approximately 4 kcal of food energy per gram as do other digestible sugars and starches. It is therefore important that diabetics understand that it is a sugar and for everyone to understand that it contributes Calories. The enzyme trehalase is found in human intestinal system and kidneys although its activity varies and it may be missing in some people. In those cases, malabsorption similar to that found with consumption of lactose in lactose intolerant individuals, or sugar alcohols in most people, may be observed. Fewer persons are thought to be trehalase deficient than are lactase deficient.”
Like humans, mice have trehalase in their guts, developing its expression early in their lives (ref)(ref)(ref). The experiments described above based on adding trehalose to mouse drinking water suggest that significant benefits are nonetheless realized via upgrading autophagy and mitigating insulin resistance(ref)(ref)(ref). So, some trehalose must be getting into cells. I infer this is due to a certain portion of the trehhalose evading gut trehalase breakdown and passing into the bloodstream where it is later broken down by circulating trehalase or in kidneys. I further infer that that the same happens when humans consume trehalose. The percentage amount entering the bloodstream is unknown. However as identified above, it is thought that 5 gms per day is a safe amount for human consumption.
12. Not all sugars are equally healthy. Fructose, in contrast to trehalose, triggers pro-aging and procancer IGF-1 signaling, mTOR activation and inhibition of FOXO3A.
Fructose appears to be a particularly evil sugar. It does this by binding to the GLUT5 transporter. Here is a rather complex diagram on carbohydrate digestion and absorption applicable to humans as well as the naked mole rat.
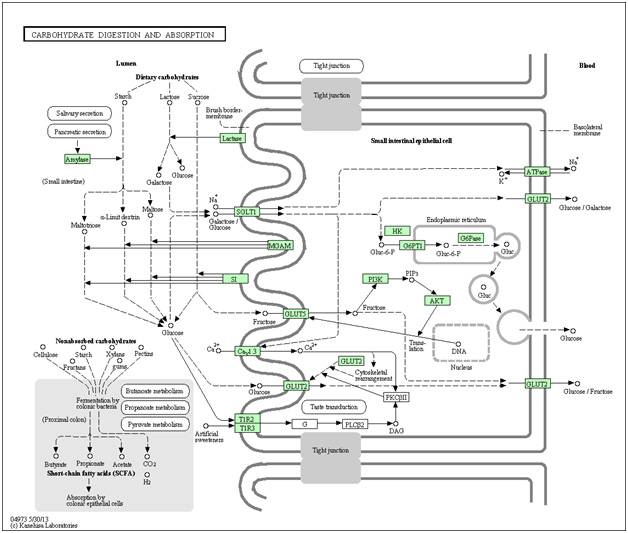
Image and legend source “Dietary carbohydrate in humans and omnivorous animals is a major nutrient. The carbohydrates that we ingest vary from the lactose in milk to complex carbohydrates. These carbohydrates are digested to monosaccharides, mostly glucose, galactose and fructose, prior to absorption in the small intestine. Glucose and galactose are initially transported into the enterocyte by SGLT1 located in the apical brush border membrane and then exit through the basolateral membrane by either GLUT2 or exocytosis. In a new model of intestinal glucose absorption, transport by SGLT1 induces rapid insertion and activation of GLUT2 in the brush border membrane by a PKC betaII-dependent mechanism. Moreover, trafficking of apical GLUT2 is rapidly up-regulated by glucose and artificial sweeteners, which act through T1R2 + T1R3/alpha-gustducin to activate PLC-beta2 and PKC-beta II. Fructose is transported separately by the brush border GLUT5 and then released out of the enterocyte into the blood by GLUT2.”
Interpretation: fructose is the only sugar absorber by the GLUT5 transporter. Once in the cell fructose directly activates the intracellular portion if the Insulin/IGF-1 pathway (PI3K/PIP3/Akt kinase cascade). The net result of Akt activation is mTOR activation and inhibition of the FOXO transcription factors, of which FOXO3a is a bona fide longevity gene. This is why high fructose corn syrup probably shortens lifespan and decreases health span. It is worse than glucose.
Driving through Iowa a few years back I noticed long train after long train of tank cars and vast sidings loaded with them – all for high fructose corn syrup, not petroleum. Among the hundreds or thousands of processed foods and medicines containing this fructose syrup are the ones on this list.
13. There are issues with using animal knockout models that could lead to lack of translatability of some of the above findings to aging humans
Most of the studies on the beneficial effects of trehalose on protein aggregations in neurodegenerative diseases listed above have been have been based on use of transgenic animal models. That is, on use of probably-younger mice where critical genes have been knocked out so as to lead to an animal model of the human disease to be simulated, be it Huntingtons, Alzheimers, etc. The main beneficial effects seem to be due to induction of autophagy. As far as we can tell, these studies were not specifically on older mice. Maria Cuervo at the Albert Einstein College of Medicine is among those who have written extensively about how aging impedes autophagy by inducing changes in cell lipid membranes. Why chaperone-mediated autophagy tends not to work with aging is because of age-related contamination of liposomal membranes. Therefore, trehalose may not work or work less well to induce autophagy in older animals or people. See Altered dynamics of the lysosomal receptor for chaperone-mediated autophagy with age. “We conclude that age-related changes at the lysosomal membrane are responsible for the altered turnover of the CMA receptor in old organisms and the consequent decline in this pathway.” This age-related effect tcould be due, for example, to consumption of a high fat or cholesterol-rich diet(ref)
On the other hand, there are publications that suggests that age-related decline in autophagy in older mice can be restored via trehalose: The 2012 publication Translational evidence that impaired autophagy contributes to arterial ageing relates: “In old mice, treatment with the autophagy-enhancing agent trehalose restored expression of autophagy markers, rescued NO-mediated EDD by reducing oxidative stress, and normalized inflammatory cytokine expression. In cultured endothelial cells, inhibition of autophagy increased oxidative stress and reduced NO production, whereas trehalose enhanced NO production via an autophagy-dependent mechanism. These results provide the first evidence that autophagy is impaired with ageing in vascular tissues. Our findings also suggest that autophagy preserves arterial endothelial function by reducing oxidative stress and inflammation and increasing NO bioavailability. Autophagy-enhancing therapies may therefore have therapeutic efficacy for ameliorating age-associated arterial dysfunction and preventing CVD.” Also spermadine is among other autophagy-inducers that can reverse arterial aging(ref).
So, it appears to me (Vince) that the bottom line on whether some or all of the beneficial autophshy and IGF-1 suppression effects of trehalose observed in mouse models translate to aged humans is not in.
14. Supplementation with trehalose could lead to people with a rare disease, trehalase deficiency disorder, to have problems.
“Trehalase deficiency is a metabolic condition in which the body is not able to convert trehalose, a disaccharide found in mushrooms, algae and insects, into glucose.[1][2][3] Individuals with this deficiency experience vomiting, abdominal discomfort and diarrhea after eating foods containing trehalose.[4][5] Studies suggest that this condition is very rare in white Americans, very frequent in Greenland natives (where it has been reported in 8% of the population), and not infrequent among Finns.[2][3][4][6] However, since ingestion of large quantities of foods containing trehalose in not common, the real frequency in unknown. Most cases appear to be inherited in an autosomal recessive manner.[2](ref)”
15. Scientific studies clearly establishing health benefits of trehalose supplementation in humans appear to be conspicuously missing
I am frankly puzzled and bothered by the fact that as far as I (Vince) can tell, there are no human studies related to the health and longevity impacts of trehalose other than for the old safety studies and the few cell-level studies mentioned above. So we don’t know whether health benefits can actually be achieved by human consumption of trehalose. One would think that the string of animal studies listed above showing very positive health impacts based on understandable mechanisms of action would lead to a big impetus to see if the same results hold up in humans – and to see whether trehalose can help us with our otherwise intractable neurological and metabolic diseases. And, possibly help is to live longer. But I found no published clinical trials, no population studies and not even any tiny studies testing the consequences of trehelulose consumption by a few people. Note that all the studies I listed above where published in past years. I can understand why drug companies will not invest money in a natural sugar that can’t be patented. But what about university researchers and public health advocates? If it does such great things for mouse health when added to their drinking water, shouldn’t we be thinking of adding it to our water?
I did a search in clinicaltrials,gov using the term “trehalose” and was at first very excited to retrieve 1023 studies in various stages of competion. A closer look, however, revealed that almost all of these retrieved studies are not really about trehalose (e.g. the studies were retrieved because trehalose plays a role as a cancer biomarker) and only 2 or 3 of the studies are actually about testing health benefits of trehalose in humans. One trial in particular, might get the ball rolling. Oral Trehalose Therapy to Reverse Arterial Aging in Middle-Aged and Older Adults, sponsored by the University of Colorado in Boulder. The trial was initiated in 2012 and is still in the recruiting stage. “The proposed study will assess the ability of a naturally occurring sugar to improve the function of arteries with age. Overall, the proposed research project has the long-term potential to influence clinical practice guidelines by establishing a novel, easy to deliver, cost-effective therapy for treating age-associated arterial dysfunction and reducing the risk of cardiovascular disease with age.”
So, is trehalose a sugar mainly interesting because it is absorbed slowly and therefore less likely to initiate sudden glucose level spikes? Or is trehalose a powerful chaperone and autophagy-inducing substance that could be beneficial for the prevention and treatment of major neurological disease in humans? Can consumption of trehalose slow human aging? We don’t know and the way things are going it looks like it it will be a long time before we find out. The research required to answer these questions mostly does not seem to be going on.

http://www.fasebj.org/cgi/content/meeting_abstract/25/1_MeetingAbstracts/lb466
This may be the reason why clinical trial is being conducted on the arterial aging of humans. This is already proved in rats.
I enjoyed this article, yet the findings puzzled me as much as they did you. On the one hand the ingestion of trehalose, at least in other species, appears to produce health and longevity benefits. Yet trehalase deficiency is harmful, and trehalase breaks down trehalose into glucose before ingestion, which would make it appear that trehalose ingestion is tantamount to ingesting an equivalent amount of glucose. You wonder whether a small amount of trehalose that bypasses trehalase enzyme might be entering the body and at a small dose causing positive hormetic? effects?
I thought of another possibility which might be testable. Could ingestion of trehalose have some effect on the intestinal flora, resulting indirectly in health benefits? Perhaps trehalose affects bacteria differently than glucose, and some is ingested by these bacterial flora. If so, this would mean that no trehalose would necessarily have to enter the body to cause health effects. for that matter, we could also ask the same question of any supplements we take. it is already thought that the types of flora in our guts affect the development of obesity and possibly other diseases.
I have ingested trehalose sugar at a rate of over 100 pounds per year for more than 2 years without side affects. You are welcome to contact me and discuss my results.
I am very interested in hearing about what positive results you have experienced with trehalose. Thank you in advance for your response.
I have been eating small doses 10 gs a day (no noticeable effects)…I’m curious how one could get up to ~150 grams a day both financially or practically…what would the recipe(s) or solution be? As William asks what are the results? I recently bought a bulk bag of Trehalose off ebay…My intention is to up my carbs with Trehalose following a cyclical Carb-back-loading protocol. I’ll follow up with any significant changes myself.
Just to follow up:
Either I have a bad batch of Trehalose with a concomitant irritant or I am not a producer of Trehalase. I say this because at about 10grams/day I have noticeably elevated HR (55ish->85ish), BP elevation, and gas. With higher doses I have pain presumably from gas. I’m going to abstain from this intervention perhaps forever. I am scotch-english ancestry so perhaps I overlap genetically with the Icelanders and I don’t produce the enzyme? The problems only came with the bulk bag…so I might retest the Swanson brand in some time. I’d like to see more compelling literature before I extend the effort though.
I have PD and have just started eating trehalose to the tune of 10 grams per day. So far I have not had any observable side effects. Mine came from Swanson.
Pingback: Fight The Harmful Effects Of Sugar With This Natural Sugar And Prevent Fatty Liver Disease - Juicing for Health